The last few years have seen several significant finds related to human prehistory. There was the 2003 discovery of Homo floresiensis, aka “the hobbit”, a distinct (and extinct) species in the same genus as us, Homo. In 2017, more accurate dating pushed back the date of human arrival to Australia by ~25,000 years, therefore increasing the estimated overlap between humans and “hobbits” by the same time span. And just a couple of months ago a paper announced that Homo sapiens bones found in Morocco had been dated from 300,000 years ago (ya), pushing back our species’ origin by about 100,000 years, and calling into question some details around the East Africa origin theory.
Technology advances, particularly in the fields of dating and DNA analysis are greatly increasing our understanding of hominid evolutionary history. These are exciting times if you’re interested in this field. In this two-part article I will provide an overview of the current state of knowledge, and touch on key points of interest. I must make an important disclaimer – the study of human prehistory is rife with academic disagreement and debate. I will aim to present theories that are generally agreed upon, plus alternatives where appropriate.
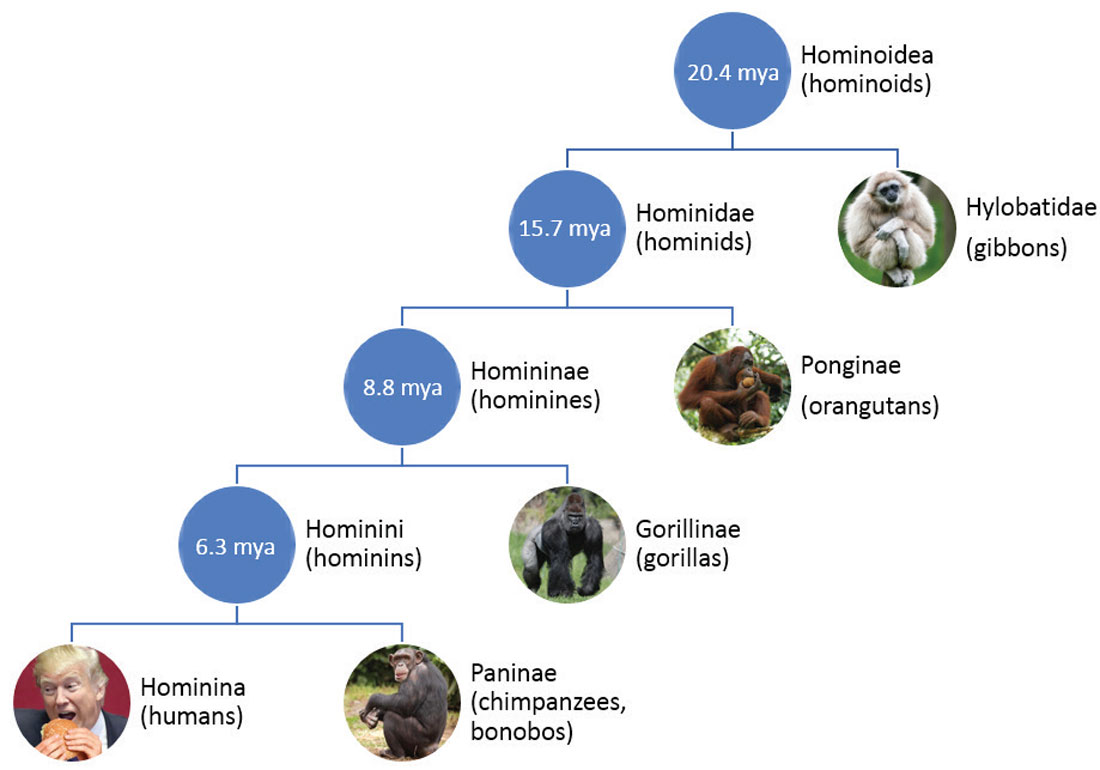
Figure 1 shows the taxonomy of the Hominoidea superfamily tree, which contains all known apes. Hominoidea split about 20.4 million years ago (mya) into the families Hylobatidae, of which only the gibbons survive, and Hominidae, which contains the rest of the great apes, including humans. This article focuses on our genus Hominina (also referred to as Homo) after the split from the chimpanzees which occurred about 6.3 million years ago.
Figure 2 is a timeline showing significant Hominina species which evolved since the split from Paninae (or paninis as I like to call them). Recently scientists have chosen the term Hominin to refer to all Hominina species, even though that goes against traditional nomenclature rules – it’s an area of at times acrimonious taxonomic debate. I find the linguistic irony so appealing – to give the only speaking ape, Hominin, a name that sounds almost identical to the word homonym – that I have become an active supporter of the term!
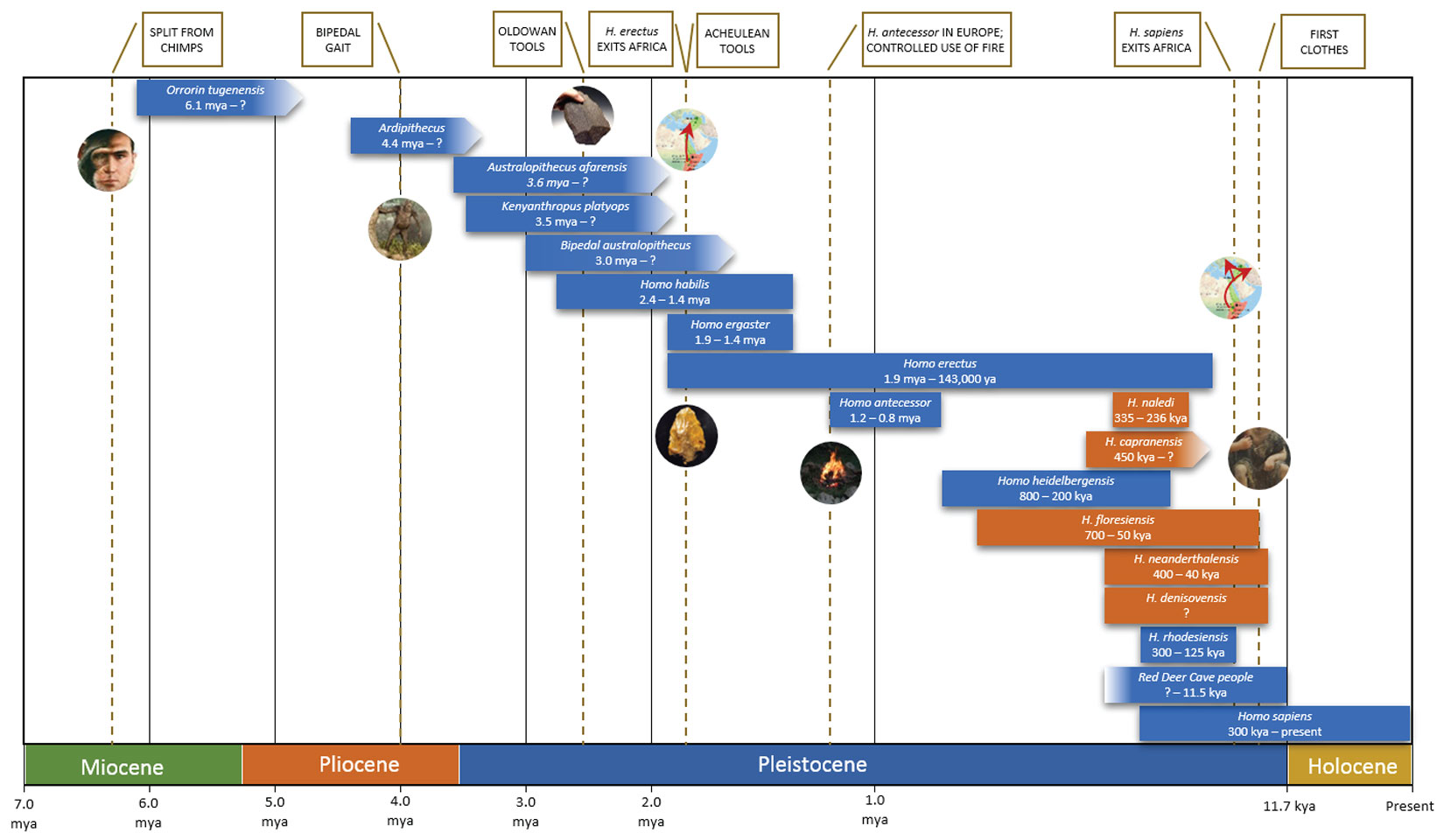
Figure 2 represents my best effort to capture the key aspects of Hominina evolution in one easy-to-understand graphic. It focuses on likely human ancestors, plus some interesting and now extinct “cousins”. To create enough room I stretched the time scale from 1 mya to the end of the Pleistocene, and stretched the Holocene even more. Vertical dashed lines indicate approximate occurrences of key advances, such as the first use of fire (defined by dating charcoal found in arranged rock hearths) and clothes (indicated by first occurrence of lice). The start and end dates for species are highly conjectural, and subject to change. It’s an exciting field – one fossil can completely blow current theories to pieces.
The fossil record shows that for most of the timeline’s 6+ million years, several Hominina species roamed the earth; our current solitary Hominina status is anomalous. For example, 200,000 years ago humans genetically the same as us shared the planet with at least nine other Hominina species. Let’s now look at some of those departed relatives.
Chimpanzee-Human Last Common Ancestor (CHLCA)
The term split suggests a sharp break, with paninis going one way, homonyms the other. In fact, the speciation process was gradual and blurry, starting as long ago as 13 million years, and continuing to as recently as 4 mya. DNA analysis suggests sporadic periods of hybridization where the two almost distinct genera hooked up again in isolated pockets – evolutionary booty calls – creating temporary periods of genetic re-convergence against a long-term trend of divergence. Evidence for this gradual and sporadic speciation comes from estimated split dates for different chromosomes; for example, geneticists estimate the two genera’s X chromosomes split 4 mya. The 6.3 mya date is thus somewhat arbitrary, but the fossil record begins to show the first human-like species right after that.
Orrorin Tugensis
This is the earliest (6.1 mya) distinctly humanoid species identified to have existed following the CHLCA split. It has been defined on the basis of 20 fossils found in the Tugen Hills, located near Lake Baringo, about 100 km north of Nakuru, Kenya. Its femur, neck and hips suggest it was somewhat bipedal, but the rest of the skeleton has features indicative of an arboreal habitat; this makes sense given its
evolutionary proximity to the CHLCA. Besides being the first clear Hominin, it is central to the debate around whether Australopithecus is a direct Homo sapiens ancestor, or part of a dead-end Hominin line. O. tugensis preceded Australopithecus by ~2m years, yet its femurs appear more human than those of Australopithecus. The debate over which of these two was our direct ancestor is ongoing (and maybe both were).
Ardipithecus
Anthropologists have identified two distinct Ardipithecus species, A. ramidus (4.4 mya) and A. kadabba (5.6 mya) based on many bones found mainly in the central Ethiopian Afar region. Arguments whether this species is part of the human lineage or not are based mainly on comparative tooth and bone morphology. It is possible that it evolved as a separate species prior to the arrival of, and distinct from, the CHLCA. Alternatively, it could have evolved later from the CHLCA, much in the same way as O. tugensis is believed to have.
Arguments for Ardipithecus being part of the human lineage are interesting, if perhaps a bit of a stretch. It possessed human-like hand features not seen in other early Hominins other than Australopithecus, and males lacked the prognathic (more protruding) jaws and larger teeth seen in chimps, baboons and other more ape-like creatures, both current and extinct. Some anthropologists link these morphological changes with human characteristics such as increased vocal ability, cooperative hunting, and generally more sophisticated social structures. They suggest that reduced sexual dimorphism indicates that Ardipithecus male status/rank was now based more on social skills, and less on brute physical dominance. In other words, Ardipithecus could represent the first step towards today’s suave male Homo sapiens, whose wit, charm, modesty and good looks are so irresistible to the female of the species.
Australopithecus Afarensis
This species was made famous by the 1974 discovery of the almost intact skeleton “Lucy” (3.2 mya), in the same Afar region where Ardipithecus lived. However, the type fossil for the species was discovered several years earlier in Laetoli, Tanzania. The significance of Au. afarensis in the human story is that it clearly establishes bipedalism. Arched feet, almost human-like hips, femurs that angle inward to centrally positioned feet, and strongly developed bone attachments for knee extensor muscles all clearly indicate this creature spent much of its time upright and walking. It did however still possess the long arms of a tree dweller, and the wrist-locking mechanism of a knuckle-walker.
In addition to bones, clear bipedal tracks were later found at Laetoli (Fig. 3). There is a strong argument that vertical stance bipedalism evolved in the trees, as gorillas and orangutans routinely walk upright on smaller, less stable branches, using their long arms for stability and additional support. Once on the ground however, they tend to knuckle walk. Additionally, or alternatively, climate changes in East Africa meant that in many places forest had given way to large patches of savannah, so perhaps this drove the development of bipedalism, with features still useful in the trees being retained. Regardless, it is easy to imagine that Au. afarensis was comfortable both on the ground and in the trees.

Kenyanthropus Platyops
Kenyanthropus platyops provides an excellent opportunity to comment on the confusion that the many Hominin species and even new genera defined by paleoanthropologists have created. The K. platyops fossils were discovered near Lake Turkana in northern Kenya by Maeve Leakey, a highly respected scientist. She is the daughter-in-law of Louis Leakey, a giant in the field whose work at Olduvai gorge over many decades really thrust our ancient relatives into public awareness. I had the good fortune to attend a primary school in Nairobi just down the road from the Kenya National Museum, where Louis Leakey was headquartered during the years when he was actively working at Olduvai and other sites. He spoke a couple of times at our school and got involved when we visited the museum; I have seen and touched many of the type fossils. Not surprisingly, I’m a lifelong paleoanthropology junkie.
Maeve Leakey has proposed that K. platyops represents a distinct species, and possibly genus. Similarly, other scientists have identified their fossil finds as totally distinct. What always follows is a hue and cry that these are merely variations within clearly established species or genera. Given that often these arguments are based on a handful of contentiously dated bone fragments, totally divergent opinions can arise, and obviously we lack sufficient data points to conclusively resolve most of these questions. However, each new find adds more information and our core understandings come into better focus. In this spirit I have left numerous possible species out of this overview; for example, rather than delving into the many possible Australopithecine species, I have focused on Au. afarensis.
Some consider Kenyanthropus platyops to be a type of Au. afarensis, but if it was not, then it was contemporary, and many experts believe K. platyops to be our direct ancestor, not Au. afarensis. This is largely because the former possessed the same flat face and small molars that we have. I often wonder why convergent evolution is downplayed or not mentioned at all in discussions such as these. Examples of this are all around us – alligators/crocodiles, bats/birds, etc. – yet anthropologists routinely place ancient species in the human lineage based on things like similarity of molars. Could it not be equally possible that K. platyops is not related to humans and just developed molars like ours because of a similar omnivorous diet?
Homo Habilis
In 1960 Louis Leakey discovered the Homo habilis type specimen in Olduvai Gorge. Its Latin name translates to ‘handy man’, so given because at the time it was believed to have been the first hominid to master the use of stone hand tools. Its taxonomy and relationship to modern humans has been hotly debated ever since its discovery. Some believe it to be a distinct species that derived from Au. afarensis, and subsequently evolved into Homo erectus. A jaw bone discovered in Afar in 2013 and dated to 2.8 mya appears to support this theory, as it morphologically appears to be half way between an Au. afarensis jaw and a H. habilis jaw. A variation on this theory is that H. habilis evolved into H. ergaster, which then evolved into H. erectus.
Alternatively, H. habilis could be a variety of Au. afarensis, or merely a combination of misidentified Au. afarensis and H. erectus bones. Recent finds in Ileret and Lake Turkana, northern Kenya, suggest that H. habilis and H. erectus coexisted, and that they could be distinct species descended from Au. afarensis, or that H. erectus had a cladogenic relationship to H. habilis, meaning that an isolated sub-group of H. habilis evolved into H. erectus while the main population of H. habilis continued, relatively unchanged. Regardless, there is a broad agreement that H. habilis is the first clear member of the genus Homo. H. habilis is known for the development and use of Olduwan hand axe technology, and many contemporaneous animal bones show signs of being butchered with these primitive hand tools.
Homo Paranthropus
I would be remiss to not mention Homo paranthropus. Back in the Louis Leakey era, this genus was central to heated debates, mainly around where it fit into the larger Homo story, especially whether it was part of our human lineage. That debate has largely passed, and H. paranthropus has been sidelined to a genus encompassing several species contemporary with H. habilis and H. erectus, which eventually went extinct. Their fossils are often found in the same sites as these other species, and this presented many challenges in the early days of discovery. However, once their distinct morphologies were established, it was clear they represented a different genus.
Paranthropus was similar to H. habilis, but stockier and more robust, with a relatively massive jaw. Its habitat was likely more in the woodlands than the savannah favoured by H. habilis, and it probably remained restricted to its preferred diet of plants and grubs, as opposed to H. habilis which is assumed to have moved towards a broader, more omnivorous diet. Lasting approximately 1.5 million years, hardly an evolutionary failure, paranthropus is often narcissistically described as a bit of a loser, because it didn’t have the good fortune to be part our “winning” human lineage. It probably did die out eventually due to climate change, whereas the line following H. habilis survived, being more environmentally flexible creatures.
Homo Ergaster
Homo ergaster is believed to have evolved from H. habilis. As you might by now predict, there is also controversy surrounding Homo ergaster’s place in the Hominin family tree, specifically its relation to Homo erectus. Was it an ancestor of H. erectus or a cousin? Or are the H. ergaster bones just creatively misidentified H. erectus bones? Many anthropologists believe that H. ergaster represents an East and South African population that had diverged sufficiently from H. erectus to be considered a separate species. Others believe H. ergaster or a “paleo-erectus” emigrated out of Africa and evolved into H. erectus in its non-African territory, leaving behind H. ergaster to evolve separately into a strictly African species.
It is clear that H. erectus was the first Hominin known to have expanded out of Africa, and the wide variation in the size and shape of H. erectus skulls recently found in Dmanisi, Georgia suggest many or all of the Homo species identified to have existed in Africa around the time of H. erectus, including H. habilis, H. ergaster, H. rudolfiensis and H. erectus itself, are all one and the same species, and just represent variations. It reminds me of something my father used to say: “A language is a dialect with an army to back it up.” Similarly, in the early Homo landscape, it could be said that a species is a variation backed by scientists with sufficient academic clout. Often a type of national or regional pride is at play, where each region’s paleoanthropologists believe their fossils are a unique species. Until I see indisputable evidence to the contrary, I am going to view Homo erectus as including variants such as H. ergaster.
Homo Erectus
Leaving behind the muddle that was Hominin East Africa from ~2.5 to 1.5 mya, let’s look at Homo erectus, which famously left Africa and successfully spread into Europe and across Eurasia all the way to S.E. Asia. It first shows up in the fossil record around 1.9 mya, was established outside of Africa “just” 100,000 years later, and existed until 143,000 ya in Java; this definitely represents the record for longevity among Hominin species. Presumably H. erectus evolved directly from some Australopithecine species, Ardipithecus, or some intermediary species including some of the ones I’ve mentioned above.
Beginning in 1891 in Java, Indonesia, most of the early H. erectus finds were in Asia, including the famous Peking man found in Zhoukoudian, China in 1921. Remember that Darwin’s “On the Origin of the Species” had just recently been published in 1859, and the scientific community was still very actively digesting its implications, and conducting searches for ancient species, including humanoid, and constructing and debating various evolutionary family trees. The popular thinking at this time was that humans had evolved in Asia and spread outwards from there; in other words, back to Africa. Eventually the large number of more primitive H. erectus fossils found in Africa swung the consensus view 180 degrees to the out of Africa theory, which contends that humans originated in Africa.
Early H. erectus skulls, such as those found in Georgia and dated to 1.8 mya, can be quite small, with a cranial volume sometimes less than 600 cm3, while later ones such as those found in Java can be human sized, up to 1100 cm3 (modern humans average 1130 for females, 1260 for males). Given the length of time H. erectus survived as a species, it is not surprising to see significant changes. Besides being viewed as an ancestor of ours, H. erectus spawned several other Hominin species such as H. neanderthalensis, H. heidelbergensis, and H. denisovensis, some of which probably interbred with early H. sapiens (more on that in part 2). It’s most likely that we, Homo sapiens, descended in a direct line from H. erectus in Africa, with some earlier outside-of-Africa offshoots contributing to our lineage at a later date.
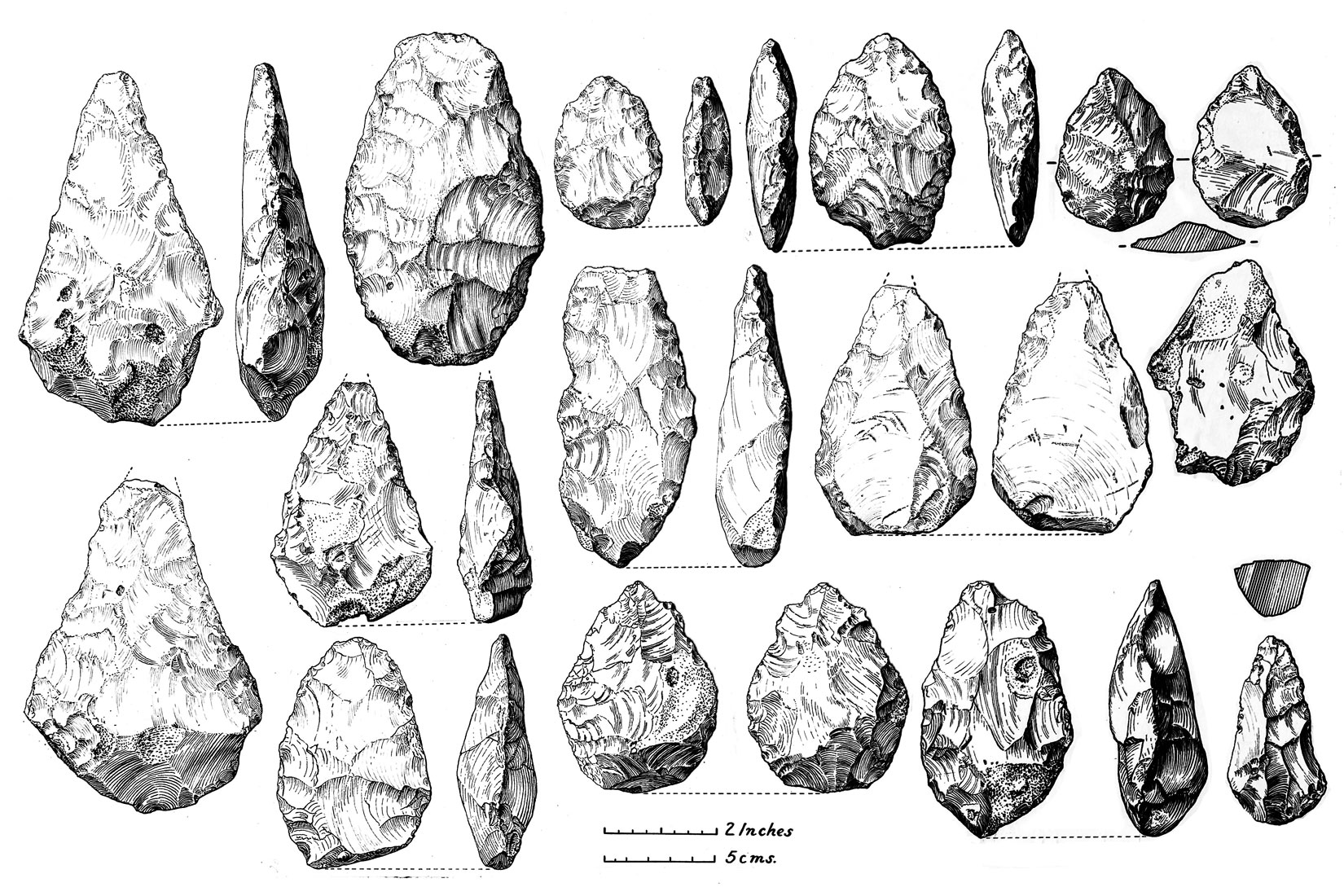
Homo erectus inherited the Olduwan tool set used by H. habilis and refined it to produce the classic Acheulean tools (Figure 4). This is just one of several technical advances that H. erectus introduced and which helped make it such a widespread and successful species. They were most likely the first species to use fire in a controlled way, and probably had to master the construction and use of boats (to reach places that were islands even during ice ages when sea levels were lower). Increased brain size, more social cooperation, apparent care of the aged and infirm, and the development of more complex language skills most likely accompanied these technical advances and resulted in a hunter gatherer lifestyle that put H. erectus in good stead for almost two million years. We can still see vestiges of that lifestyle in some modern human tribes, and if there ever is some kind of apocalyptic breakdown of modern society, it will be those humans who survive, not us cellphone-toting weaklings.
Homo Antecessor
In addition to spreading east, H. erectus most likely also pushed up into Europe, evidenced by stone axes found in Turkey, along the way, so to speak. One way or another, Hominins started to appear there around 1.2 mya in the form of Homo antecessor. Either H. erectus got there first and evolved into H. antecessor, or H. antecessor evolved separately in Africa and then entered Europe. Regardless, fossils found at two sites in Spain have been classified as Homo antecessor, the first Hominin known to have lived in Europe. There are hints that they may have ranged as far north as France and England – footprints exposed on a Norfolk beach and dated at 850,000 years old were most likely left by a H. antecessor individual.
Homo Heidelbergensis
The common belief is that Homo heidelbergensis evolved in Africa, and then migrated north into Europe and the Middle East. The main debate surrounding H. heidelbergensis is whether or not it qualifies to be a separate species that evolved from H. erectus. The alternative view is that H. erectus gradually morphed over time into what has been identified as H. heidelbergensis, and this gradual change continued to produce Homo neanderthalensis and other species. The only definitive morphological change is that the cranial capacity seen in H. heidelbergensis increased by about 10% from that of H. erectus, to an average of 1,250 cm3. Fossil remains are found from South Africa, all the way to England and Germany, where of course the first fossils (but not the oldest) were found, near Heidelberg.
There are hints at H. heidelbergensis sites of familiar human habits and characteristics, such as burial of the dead, use of ochre for body paint and ritual purposes, development of spear points, and a little detail I find interesting – asymmetric tooth wear suggests these Hominini were predominantly right-handed, as we are.
There are hints at H. heidelbergensis sites of familiar human habits and characteristics, such as burial of the dead, use of ochre for body paint and ritual purposes, development of spear points, and a little detail I find interesting – asymmetric tooth wear suggests these Hominini were predominantly right-handed, as we are.
Recent developments in DNA sequencing have begun to shed light on the Hominin story from this point on. H. heidelbergensis is likely the last common ancestor shared by us (Homo sapiens) and Homo neanderthalensis, and that split would have occurred ~400,000 ya. A chronological summary for H. heidelbergensis would be (1) evolution from H. erectus, in Africa around 800,000 to 700,000 years ago; (2) movement into the Middle East and Europe around 400,000 to 300,000 years ago, and ultimate evolution into H. neanderthalensis; (3) also around 400,000 to 300,000 years ago, movement into Asia proper, and evolution into H. denisovensis; (4) evolution of the remaining African population into H. sapiens, around 300,000 years ago.
Summary
To wrap up part 1 of this article I will describe how I keep all the confusing details around early Homo evolution simple and straight in my head. I categorize the different species and variations into 5 main categories, and don’t bother too much about distinctions within each cluster.
The split from chimpanzees occurred around 6.3 mya, give or take several hundred thousand years. Immediately following this there was a cluster of species with the first hints of distinctly Homo features, such as crude bipedalism, more dexterous hands, flatter faces, smaller teeth, and larger skulls (albeit still quite small). This group includes Orrorin tugensis and Ardipithicus and they were mainly centered around Africa/Ethiopia.
The next two groups are the genera Australopithecus and Paranthropus. Paranthropus was robust, more ape-like and spanned the period between 2.7 and 1.2 mya. Australopithecus was more gracile and suited to bipedalism. Between about 3.6 to 1.5 mya it was found in a broad arch with one end in South Africa/Namibia, that curved through East Africa/Ethiopia and into Chad and ended in what is now the middle of the Sahara Desert.
The fourth major group is the genus Homo, which we are the last remaining member of. A key step for this group was the expansion out of Africa by H. erectus just under 2 mya. Other species/variations in this group include H. antecessor and H. heidelbergensis. H. heidelbergensis then directly and indirectly spawned several subsequent Homo species, which together represent the 5th and last cluster, to be covered in part 2.
Most scientific debates until recently have been morphological and involved taxonomic questions such as whether certain fossils represent a different genus or species, and how different species were related to each other, especially whether they were part of the direct human lineage. I’m not sure all that matters much, because the data points are few and far between. But each new find helps fill in the gaps a bit more. What is really exciting lately is that DNA evidence is now providing more quantitative information on these questions. It is unlikely that there really is a clearly defined lineage to be discovered; recent DNA analysis shows how complex a species’ evolution can be. Sub-populations can diverge in isolation and converge again with the main group, often, and in different places and times. That is most likely was going on in Africa during the early Hominin days. Let’s face it, us apes have healthy libidos and the various early Homo species were surely no different, and lots of “cross-pollination” was surely going on.
In the second part of this article we’ll pick up the story from H. heidelbergensis, look at the various species that followed it, and delve into DNA sequencing and how it is illuminating the relationships between these more recent Homo species.








Share This Column